| Ischigualasto Formation | |
|---|---|
| Stratigraphic range: Late Carnian - Early Norian | |
 Valle Pintado, Ischigualasto Formation | |
| Type | Geological formation |
| Unit of | Agua de la Peña Group |
| Sub-units | Quebrada de la Sal, Valle de la Luna, Cancha de Bochas & La Peña Members |
| Underlies | Los Colorados Formation |
| Overlies | Los Rastros Formation |
| Thickness | Up to 1,059 m (3,474 ft) |
| Lithology | |
| Primary | Sandstone, mudstone |
| Other | Tuff, conglomerate, siltstone, basalt |
| Location | |
| Coordinates | 29°36′S 68°06′W / 29.6°S 68.1°W |
| Approximate paleocoordinates | 46°00′S 40°12′W / 46.0°S 40.2°W |
| Region | La Rioja Province & San Juan Province |
| Country | Argentina |
| Extent | Ischigualasto-Villa Unión Basin |
| Type section | |
| Named for | Cacán: "Place where the moon alights" |
| Named by | Frenguelli (1944) |
 Ischigualasto Formation (Argentina) | |
The Ischigualasto Formation is a Late Triassic fossiliferous formation and konzentrat-lagerstätte in the Ischigualasto-Villa Unión Basin of the southwestern La Rioja Province and northeastern San Juan Province in northwestern Argentina. The formation dates to the Carnian and Norian ages and ranges between 231.7 and 225 Ma, based on ash bed dating.
The up to 900 metres (3,000 ft) thick formation is part of the Agua de la Peña Group, overlies Los Rastros Formation and is overlain by Los Colorados Formation. The formation is subdivided into four members, from old to young; La Peña, Cancha de Bochas, Valle de la Luna and Quebrada de la Sal. The sandstones, mudstones, conglomerates and tuffs of the formation were deposited in a humid alluvial to fluvial floodplain environment, characterized by strongly seasonal rainfall.
The Ischigualasto Formation is an important paleontological unit and considered a Lagerstätte, as it preserves several genera of early dinosaurs, other archosaurs, synapsids, and temnospondyls of the Late Triassic. Coprolites and fossil wood also have been found in the formation. The formation crops out in the 1967 established Ischigualasto Provincial Park, which was designated a UNESCO World Heritage Site in 2000.
Etymology
The name Ischigualasto is derived from the extinct Cacán language, spoken by an indigenous group referred to as the Diaguita by the Spanish conquistadors and means "place where the moon alights".[1] The genus Ischigualastia and the species Herrerasaurus ischigualastensis, Pseudochampsa ischigualastensis, Pelorocephalus ischigualastensis and Protojuniperoxylon ischigualastianus were named after the formation.
Geology
Regional setting
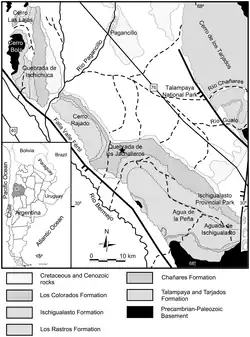
The Ischigualasto Formation is found in the Ischigualasto-Villa Unión Basin, along the border between La Rioja and San Juan Provinces in the badlands of Western Argentina. The formation is most widely exposed and studied in Ischigualasto Provincial Park (IPP), an Argentine national park which occupies a large portion of the basin in San Juan province. In the western part of the park, the maximum thickness of the formation amounts to 691 metres (2,267 ft), narrowing to 397 metres (1,302 ft) in the eastern area.[2][3] Conversely, the formation is thin and mostly covered by recent sediments in Talampaya National Park, which neighbors Ischigualasto Provincial Park in La Rioja to the north.[4] Not every exposure in La Rioja is so limited, however. The previously-neglected Cerro Las Lajas area, at the northwest tip of the basin, preserves a 1,059-metre (3,474-foot) thick outcrop in a small geographic area.[3] Another well-exposed outcrop in La Rioja is Cerro Bola, which preserves nearly 800 metres (2,600 feet) of sediment.[5]
The Ischigualasto-Villa Unión Basin is the remnant of an ancient half-graben which preserves a 3.5-kilometre (11,000 ft)[6]-thick series of Triassic sediments.[7] The Ischigualasto Formation represents the second syn-rift period within the basin, meaning that its sediments were implaced during a brief interval when rifting was reactivated to widen the basin further. The rifting was accommodated by movement along Valle Fértil, a dormant fault at the northwest corner of the basin.[2]
Stratigraphically, the formation is part of the Agua de la Peña Group, a name encompassing Late Triassic formations within the Ischigualasto-Villa Unión Basin. The Ischigualasto Formation overlies the Los Rastros Formation, a mid-Carnian unit dominated by green mudstone. It is subsequently overlain by the Los Colorados Formation, which is primarily Norian-age red sandstone. The comparatively pale color of the Ischigualasto Formation's sediments strongly contrasts with its predecessor and successor, helping to distinguish the three formations in the field.[8]
Sedimentology
The Ischigualasto Formation was a river-dominated environment as indicated by its rock types: coarse fluvial (river channel) sandstone beds, and finer floodplain deposits of sandstones, mudstones, and paleosols.[2][9] Thin bentonite tuffs are scattered through the entire formation, derived from volcanic ash.[2][10][3]
Sandstone beds
The coarsest rocks in the formation are channel lag pebble-cobble river conglomerates (i.e., gravel beds, the first river sediments to sink and become buried). These form the lowermost layer of many sandstone beds, eroding into underlying fine-grained sediments. Flat, ribbon-like layers are the most common variety of river sandstone, and some layers preserve underwater ripples or dunes. Slanted sandstone layers indicate sediment accretion, either downstream or laterally (in the form of point bars). Sandstone beds occupy a range of sizes and proportions, from relatively small single-channel deposits (2 meters thick, 10 meters wide), up to massive complexes of multiple superimposed braided rivers (20 meters thick, 2000 meters wide).[2] Sandstone and siltstone also occur in the surrounding floodplain, in the form of crevasse splays, abandoned channels, and levee deposits.[2]
The ancient rivers and streams mainly flowed towards the north or northeast from volcanic highlands flanking the southern part of a prehistoric valley. At the southeastern edge of the modern basin, where the Ischigualasto Formation is somewhat thinner, flow indicators trend towards a northwestward orientation, possibly redirected by basalt flows. The sandstone beds also tend to be smaller and less numerous in the eastern part of the basin, while paleosols are more voluminous.[2]
Mudstones and paleosols
Most floodplain sediments were fine-grained. Many of the coarser mudstone layers are overbank deposits, laid down over abandoned channels and crevasse splays.[2] The finest sediments are preserved as paleosols (fossilized soil), of which there were eight varieties, labelled A through H.[11][2] Type A paleosols are sandy soils rich in quartz and mica, with indistinct mottling, root casts, and no internal structure. They were probably protosols or entisols, young soils developing along streambanks where siliciclastic grains outnumber organic material.[11] The other paleosols are mostly found among claystone or mudstone beds rather than sandy layers. Type B paleosols have high proportions of smectite and mica, frequent slickensides, wedge-shaped crevices (mudcracks in cross-section), and strong redoximorphic features such as gleying and mottling. They can be classified are a type of vertisol, emplaced in a warm seasonal climate with a high water table during much of the year.[11] Type C paleosols are similar to type B, yet differ in their abundance of calcareous nodules and a paucity of redoximorphic features. Though also vertisols, they correlate with a less humid climate, moistened by periodic rainfall rather than groundwater.[11]
Type D paleosols have high smectite and kaolinite, fine mottling, and distinctively numerous hematite nodules and clay films (argillic horizons). They are classified as argillisols or alfisols, buried in relatively dry and stable environments such as forests, away from the influence of water.[11] Type E paleosols are simple blocky soils packed with calcareous structures and a distinct carbonate horizon alongside smectite and mica. They are calcisols, perpetually dry and stable desert soils forming in an climate with little precipitation.[11] Type F paleosols are intermediate between type D and type E, since they have both clay- and carbonate-rich horizons and all three major clay minerals: smectite, mica, and kaolinite.[11]
Type G and H paleosols are distinct in that they are derived from volcanic rocks, rather than sediments. Type G paleosols are colorful and sandy soils with distinct horizons rich in smectite, silica, and iron oxides. They develop above feldspar-based ash beds and are among the most laterally extensive soil type in the formation, since volcanic ash covers a broad area without regard to climate or water availability.[11] Type H paleosols, on the other hand, are a rare variety of granular paleosol developing above basalt lavas which have been degraded and weathered during the process of soil formation.[11]
Subdivisions
The formation is subdivided four members in Ischigualasto Provincial Park, with each member showing a distinctive pattern of sedimentology and fossil content.[2] These members were informally delineated in 2006[11] and formally named in 2009.[2] From top (youngest and stratigraphically highest) to bottom (oldest and stratigraphically lowest), they are:[2][12]
- Quebrada de la Sal Member: ~60 metres (200 ft). A coarse-grained member nearly lacking in fossil content. Most sediments are brown or red in color, though grey or mottled mudstones still occur sporadically, unlike the overlying Los Colorados Formation. The upper extent of the member (and the Ischigualasto Formation as a whole) is defined by the last grey mudstone bed in the Ischigualasto-Villa Unión Basin. All variety of sandstone features can be found in this member.[2] Paleosols are limited and similar to those of the La Peña member, with type A and to a lesser extent type D as the most common varieties. Type B, F, and G paleosols are present but exceedingly rare.[11]
- Valle de la Luna Member: ~450 metres (1,480 ft). The thickest member in the formation. Dark grey smectite-rich floodplain mudstone is the most common lithology, especially in the eastern part of the basin. Sandstone features are common and well-developed in the western part of the basin, as are abandoned channel overbank deposits with a high proportion of calcareous sediments. Tetrapod fossils continue to decrease in prevalence relative to earlier members, but plant fossils are locally abundant in abandoned channel deposits.[2] The western basin has a high proportion of type A, D, and G paleosols, the central basin has more type B, and the eastern basin is mostly type A and B. Type C, E, and F paleosols are practically absent outside of the eastern basin.[11]
- Cancha de Bochas Member: ~130 metres (430 ft). A fine-grained member mainly composed of mottled red/green/gray mudstone. Fossils are common, though not to the same degree as the La Peña Member. Calcareous nodules are abundant and used to distinguish this member from the La Peña and Valle de la Luna members. Slickensides, root casts, and other hallmarks of soil alteration are frequently encountered as well. Both single-channel and multi-channel sandstone beds are present, as are crevasse splay and levee deposits. Volcanic rocks include not just tuff, but also a thick basalt extrusion in the eastern part of the basin.[2] This member hosts every type of paleosol except for type G. It is also the only member where type H paleosols are present. Type A is most common overall, type C is the most common in the middle, and types E, F, and B are most frequently found in the eastern area.[11]
- La Peña Member: ~50 metres (160 ft). A coarse-grained member with abundant fossils. Its base is defined by the first massive channel conglomerate layer above the Los Rastros Formation. Multi-channel fluvial sandstone is common and tannish-grey in color, though isolated single-channel and point bar deposits are seemingly absent, in contrast to the rest of the formation. Crevasse splay deposits are present but thin and silty, and floodplain mudstone is greenish-gray, sandy, and rich in smectite.[2] Paleosols are uncommon; the western basin has type A and very rare type D paleosols while the eastern basin mostly has type B.[11]
The outcrop at Cerro Las Lajas is instead divided into three subunits: a fossiliferous lower section (11 to 310 meters above the base of the outcrop) with meandering river deposits, a fossil-poor middle section (310 to 740 meters above base) with high-humidity paleosols, and an upper section (740 to 1070 meters above base) with braided river deposits and welded tuff beds, but no fossils.[3]
Age
Interlayered volcanic ash layers above the base and below the top of the Ischigualasto Formation in Ischigualasto Provincial Park provide precise chronostratigraphic control on the formation's duration. The two ash beds have yielded ages of 231.4 ± 0.3 Ma and 225.9 ± 0.9 Ma, respectively. These age estimates were derived from 40Ar/39Ar radiometric dating on sediment samples first analyzed in 1993[10] and recalibrated in 2011.[13][14] The formation as a whole spans the Carnian-Norian boundary (at approximately 227 Ma), though the greatest diversity occurs in the older Carnian-age members of the formation (the La Peña and Cancha de Bochas members).[12] Palynomorph and tetrapod biostratigraphy agree with a late Carnian to early Norian age for the formation.[15][16][3][17] A major faunal turnover and loss of diversity occurs in the early part of the Valle de la Luna Member, shortly before the start of the Norian.[12] According to U-Pb dating, a 2021 study published an age of 228.91 ± 0.14 Ma for an ash bed at this level.[18]
A 2020 study dated three tuffs at Cerro Las Lajas via U-Pb dating. The oldest tuff is at 107 meters above the base of the outcrop, shortly before the earliest fossils at the site. It has an age of 229.25 ± 0.1 Ma. The second tuff, in the middle of the fossiliferous section at 160 meters above base, is 228.97 ± 0.22 Ma. The third tuff is close to the top of the formation, 1035 meters above base with an age of 221.82 ± 0.1 Ma. These values estimate that the formation was deposited between 230.2 ± 1.9 Ma and 221.4 ± 1.2 at Cerro Las Lajas.[3]
Small stratigraphic gaps may have been present at Cerro Las Lajas, lending some uncertainty to the thickness of the exposure and the precise level of the tuff beds.[18][5] Even accounting for this possibility, deposition of the Ischigualasto Formation at Cerro Las Lajas seems to end several million years later than at Ischigualasto Provincial Park. The age estimates at either site may be erroneous, or the transition between the Ischigualasto and Los Colorados formations is asynchronous, with some areas acquiring sediments characteristic of the latter formation at an earlier time than other areas.[5][17] Likewise, sediments of the Los Rastros Formation may have been superseded later in Cerro Las Lajas relative to Ischigualasto Provincial Park.[5]
The Ischigualasto Formation is approximately coeval with the upper Santa Maria Formation of the Paraná Basin in southeastern Brazil, the Pekin Formation of the United States and the lower Maleri Formation of India.[19][20]
Climate and paleoenvironment
The Ischigualasto Formation dates to only a few million years after the Carnian Pluvial Episode (CPE), an interval of particularly warm and wet global climate in the mid-Carnian. The CPE corresponds to the underlying Los Rastros Formation.[21][22][23] Various studies have come to strongly different conclusions on the climate of the Ischigualasto Formation.[24][25]
A 2004 study evaluated the geological composition of an Ischigualasto paleosol derived from a weathered basalt flow about 45 m above the base of the formation.[26] The paleosol in question, later labelled as a type H paleosol,[11] was a vertisol (indicating strong wet and dry seasons) preserving a horizon rich in goethite (a weathering product facilitated by cool rainwater) alongside high concentrations of kaolinite and a lack of carbonate (both indicative of high moisture).[26] The estimated mean annual temperature was similar to Marion Island or southernmost Norway, as low as 3 to 9 °C (37 to 48 °F). Precipitation was comparable to a cool mediterranean climate, exceeding 750 to 1,000 millimeters per year (30 to 39 in/year) but with strong wet and dry seasons.[26]
Later studies indicate strong climate fluctuations through the formation, with some members characterized by a semi-arid climate.[24] When the same authors sampled a broader range of paleosols in 2006, their conclusions were modified.[11] δ18O values were extracted from calcarous nodules preserved in the three upper members of the formation. These values suggested mean annual temperatures below 11 °C (52 °F), only barely warmer than the previous estimate.[11] Precipitation was more erratic through the formation's history, as indicated by variation in the frequency of low-moisture (calcareous) and high-moisture (non-calcareous) paleosols. Calcareous paleosols (types C, E, and F) are common in the Cancha de Bochas and lower Valle de la Luna members, but rare or absent in the rest of the formation. This suggests that the middle part of the formation experienced a cool semi-arid climate while the early and later parts were more humid.[11]
A 2008 study compared the distribution of paleosols and plant fossils, with implications for climate change and the evolution of depositional environments within the Ischigualasto Formation.[27] The La Peña Member would have been subhumid and frequently destabilized by shifting braided rivers, leaving little room for the development of dry mature soils or forests.[27] By time of the Cancha de Bochas Member, the river systems stabilized into meandering and anastomosing forms, but the climate became much drier, so plant growth continued to be inhibited.[27] The upper Valle de la Luna Member was by far the wettest part of the formation, with a mean annual precipitation exceeding 760 millimeters per year (30 in/year). A high water table, stable river channels, and a more humid climate allowed for the development of marsh habitats, widespread deep soils, and large trees.[28] Dry conditions returned with the Quebrada de la Sal Member, with small sandy streams as the predominant depositional environment. These climate transitions were subtle, and the formation as a whole was assigned a climate with cool dry winters and hot rainy summers enhanced by the Pangean megamonsoon.[27][15][16] Warm temperate grasslands in Oklahoma and central Argentina were considered the closest modern equivalent to the Ischigualasto paleoenvironment.[27]
A 2022 study attempted to infer climate via a diverse set of geochemical proxies in paleosols.[5] For the Ischigualasto Formation as a whole, their results estimated a mean annual temperature of 9.4 to 17.5 °C (48.9 to 63.5 °F) and a mean annual precipitation of 700 to 1,400 millimeters per year (28 to 55 in/year). This is notably cooler and drier than the Los Rastros Formation. Using radiometrically-dated ash beds and the 800 m outcrop of Cerro Bola as guidelines, the study proposed the following climate trends through the formation: the climate started out as relatively warm and humid, before drying out (at around 100 m or 229.5 Ma), and eventually recovering a degree of fluctuating seasonal humidity at the close of the Carnian (at 380 m or 227.8 Ma). These fluctuations are difficult to compare with changes in the fauna and flora, though rhynchosaur abundance and pseudosuchian diversity may show a positive correlation with high humidity in the formation.[5]
Paleontological significance

The Ischigualasto Formation is highly fossiliferous and its unique paleontological characteristics made it a Lagerstätte; a stratigraphic unit containing a diverse faunal assemblage. The paleontological importance led to the establishment of the Ischigualasto Provincial Park, a UNESCO World Heritage Site since 2000.
The Ischigualasto Formation contains Late Triassic (Carnian) deposits (231.4 -225.9 million years before the present[14]), with some of the oldest known dinosaur remains, which are the world's foremost with regards to quality, number and importance.

Rhynchosaurs and cynodonts (especially rhynchosaur Hyperodapedon and cynodont Exaeretodon[14]) are by far the predominant findings among the tetrapod fossils in the park. A study from 1993 found dinosaur specimens to comprise only 6% of the total tetrapod sample;[10] subsequent discoveries increased this number to approximately 11% of all findings.[14] Carnivorous dinosaurs are the most common terrestrial carnivores of the Ischigualasto Formation, with herrerasaurids comprising 72% of all recovered terrestrial carnivores.[14] The carnivorous archosaur Herrerasaurus is the most numerous of these dinosaur fossils. Another important putative dinosaur with primitive characteristics is Eoraptor lunensis, found in Ischigualasto in the early 1990s.
Petrified tree trunks of Protojuniperoxylon ischigualastianus of more than 40 m (130 ft) tall attest to a rich vegetation at that time. Fossil ferns and horsetails have also been found in the formation.
Taphonomy
Some fossilized wood and tetrapod bones in the formation display particular styles of decay and degradation, helping to estimate the occurrence of various bacteria and fungi groups in the Triassic Period which otherwise lack fossil evidence.[29][30]
Most vertebrate fossils are recovered from floodplain deposits, while a smaller portion come from sandstone beds, though both sediment types exhibit a wide range of preservation quality. Fossils in the Cancha de Bochas Member are primarily preserved via calcite, while those in the La Peña and Valle de la Luna members show a higher proportion of hematite preservation.[31]
Plant preservation
Plant preservation in the Ischigualasto Formation can follow nine different taphonomic pathways, each associated with a particular depositional environment:[27]
- (A) Fibrous clay-filled root traces occupy smaller and more fine-grained sandstone beds, which were only exposed long enough for herbaceous plants to take hold. This type of preservation is common, especially in the La Peña Member and the middle part of the basin.[27]
- (B) Root halos, ring-shaped discolorations indicative of altered soil chemistry around root traces. They are found in red paleosols affected by microbial activity in upland environments. They occur from the Cancha de Bochas member upwards, with the thickest root halos in the Valle de la Luna Member.[27]
- (C) Silicified roots and stumps of woody shrubs are only found in thick braided channel sandstone deposits. The plants responsible for the silicified fossils would have inhabited dry sandbars and abandoned channels, only to be inundated and buried by new sediment and volcanic ash washed down by seasonal floods. This type of preservation is known from every member, but it is rare.[27]
- (D) Silicified roots and trunks of large woody trees, often standing perpendicular to the bedding plane. The trunks occupy abandoned channel deposits in the mid-upper Valle de la Luna Member, with the wood stabilized in position by interbedded layers of mudstone and sandstone. Rhexoxylon piatnitzkyi is the most common species of petrified wood in the formation.[27]
- (E) Horizontal silicified tree trunks and branches, parallel to the bedding plane. These trunks are found in exceptionally large sandstone beds in the mid-upper Valle de la Luna Member. They were probably riparian trees which were toppled, stripped of their roots, and buried during massive floods.[27]
- (F) Horizontal foliage impressions and cuticle, parallel to the bedding plane. These plant fossils are the most diverse and diagnostic in the formation, deposited in thin flattened layers of fine-grained sediment. The original environment was calm stagnant water, probably a shallow marsh occupying an abandoned channel near an active river. Only found in the mid-upper Valle de la Luna Member.[27]
- (G) Cuticle oriented oblique to the bedding plane, similar to the horizontal impressions but significantly more fragmentary. These are only found in trough cross-bedded sandstone, representing freshly detached foliage slowly settling onto a streambed by waning flood waters. Only found in the mid-upper Valle de la Luna Member.[27]
- (H) Palynomorphs (pollen, spores, and other microscopic plant fossils). Found in marsh mudstone alongside horizontal foliage impressions and cuticle.[27]
- (I) Eroded charcoal fragments, found alongside horizontal silicified trunks (in major flood deposits) or oblique cuticle (in trough cross-bedded sandstone beds).[27]
Paleobiota
Color key
|
Notes Uncertain or tentative taxa are in small text; |
Trace fossils
Coprolites were found in Valle Pintado in the upper part of the formation. Analysis of the coprolites revealed that plant remains were absent and bone material and apatite were sparse. The most likely candidate to have produced these fossil feces has been suggest as the most common reptile in the formation; Herrerasaurus.[32]
Two different morphotypes of animal burrows are found in levee and crevasse splay deposits in the Cancha de Bochas member.[33][34] One type of burrow is a horizontal mesh of tunnels and enlarged chambers which occasionally meet with vertical shafts branching off at right angles. Though the tunnel system extends up to 2 cubic meters, the tunnel diameters do not exceed 15 cm, while chambers only reach 25 cm in width. They were almost certainly created by small terrestrial vertebrates, presumably cynodonts.[33][34] The other tunnel morphotype is a calcareous web of narrow, elliptical burrows (average diameters of 7 cm) branching out from a long vertical shaft. It probably represents plant root casts enlarged further by small animals.[34]
Reptiles
Dinosaurs
The fossils of an undescribed species of theropod are present in San Juan Province.[35]
| Dinosaurs of the Ischigualasto Formation | |||||||
|---|---|---|---|---|---|---|---|
| Genus | Species | Province | Member | Biozone | Material | Notes | Image |
| Chromogisaurus[36] | C. novasi | San Juan[36] | lower Cancha de Bochas[36] | Hyperodapedon-Exaeretodon-Herrerasaurus biozone | A partial skeleton.[36][37] | A basal sauropodomorph closely related to Saturnalia, a saturnaliid (or saturnaliine guaibasaurid)[36] from Brazil. Estimated length is around 2 metres (6.6 ft). | |
| Eodromaeus[14] | E. murphi | San Juan[14] | La Peña,[14] Valle de la Luna[14] |
Hyperodapedon-Exaeretodon-Herrerasaurus biozone | A nearly complete skeleton and at least five other fossils.[14][12][37] | A small and well-preserved predatory dinosaur, often considered the oldest known theropod since its description in 2011.[14][37] It was a lightweight and long-limbed carnivore, around 1.2 meters (3.9 feet) in length, with an estimated weight of about 5 kilograms (11 pounds). |  |
| Eoraptor[38] | E. lunensis | San Juan[39] | Cancha de Bochas[12] | Hyperodapedon-Exaeretodon-Herrerasaurus biozone | A nearly complete skeleton[38][39][37] and at least seven other fossils, though only some of these are referred to the species with certainty.[39] | One of the most well-preserved and well-described Carnian-age dinosaurs. It was small generalist with bipedal habits and a presumably omnivorous diet based on the structure of its teeth. In its initial 1993 description, it was hailed as an early theropod vital for approximating the common ancestor of all dinosaurs.[38] Many studies have subsequently reclassified it as a basal sauropodomorph.[36][14][39][40][35][37] 1 meter (3.3 feet) in length, with an estimated weight of about 10 kilograms (22 pounds). |  |
| Herrerasaurus | H. ischigualastensis | San Juan | Cancha de Bochas[12] | Hyperodapedon-Exaeretodon-Herrerasaurus biozone | "Various partial skeletons, including a complete skull and mandible."[40][37] Over 50 specimens have been recorded.[12] | A herrerasaurid, among the largest and most well-described predatory dinosaurs of the Triassic, as well as the most common carnivore in the Ischigualasto Formation. The largest known specimen, previously referred to the discredited genus Frenguellisaurus, had a skull measuring 56 cm (22 in) in length and a total body length estimated at 6 meters (20 ft).[41][35] Though Herrerasaurus was the earliest dinosaur collected from the formation in the 1960s,[42] the most complete specimen, a juvenile, was only described in 1992.[41][43][44][45] |  |
| Panphagia[46] | P. protos | San Juan[46] |
lower Cancha de Bochas[12] |
Hyperodapedon-Exaeretodon-Herrerasaurus biozone | A disarticulated partial skeleton.[46][37] | A small basal sauropodomorph,[36][47] initially considered to be the oldest and basal-most member of the group when described in 2009.[46] Sometimes classified as a guaibasaurid or saturnaliid within Sauropodomopha. Estimated length is around 1.30 meters (4.3 feet).[46][35] |  |
| Sanjuansaurus[48] | S. gordilloi | San Juan[48] | upper La Peña[12] or lowermost Cancha de Bochas[48] | Hyperodapedon-Exaeretodon-Herrerasaurus biozone | A mostly-articulated partial skeleton.[48][37] | A herrerasaurid comparable in size to a medium-sized Herrerasaurus.[48] |  |
| An unnamed herrerasaurid[36] | Unnamed | Specimen MACN-PV 18649a, a partial forelimb.[36] | An undescribed species of herrerasaurid mentioned in conference abstracts in the late 2000s.[36] | ||||
Other archosaurs
| Non-dinosaur archosaurs of the Ischigualasto Formation[36][12] | |||||||
|---|---|---|---|---|---|---|---|
| Genus | Species | Province | Member | Biozone | Material | Notes | Images |
| Aetosauroides | A. scagliai | San Juan, La Rioja[3] |
Cancha de Bochas,[49][12] lower Valle de la Luna[12] |
Hyperodapedon-Exaeretodon-Herrerasaurus biozone, Hyperodapedon biozone,[3] Exaeretodon biozone[12] |
Armor, postcrania, and skull material[49][3] from at least 20 specimens,[12] though not all of these may belong to the genus.[49] | A large and fairly common basal aetosaur, one of the few known from South America.[49][50] Presumably carnivorous, unlike most members of the group. Once proposed to be synonymous with Stagonolepis.[51][52] |  |
| Indeterminate aetosaurs | Indeterminate | San Juan | Cancha de Bochas[53] | Hyperodapedon-Exaeretodon-Herrerasaurus biozone | Armor and postcrania of indeterminate basal aetosaurs,[49] including an articulated partial skeleton.[53] | Many indeterminate aetosaur fossils from the formation, such as the nomen dubium Argentinosuchus bonapartei, cannot be unambiguously referred to Aetosauroides.[49][12] | |
| Ignotosaurus[12] | I. fragilis | San Juan | lower Cancha de Bochas[12] | Hyperodapedon-Exaeretodon-Herrerasaurus biozone | A right ilium[12] | An obscure silesaurid dinosauromorph.[12] | 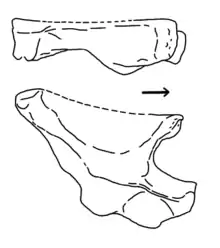 |
| Incertovenator[54] | I. longicollum | San Juan | middle Cancha de Bochas[54] | Hyperodapedon-Exaeretodon-Herrerasaurus biozone | Partial postcranial skeleton consisting of 23 articulated or semi-articulated vertebrae and a left ilium.[54] | An archosauriform of uncertain phylogenetic placement with elongated neck vertebrae. It is most parsimonously placed as an archosaur, either among Aphanosauria or as the sister taxon to Mandasuchus.[54] | 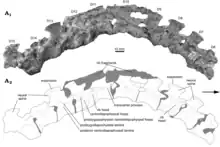 |
| Pisanosaurus[55] | P. mertii | La Rioja | Unknown ("upper levels")[12] | Hyperodapedon biozone[3] | "Fragmentary skull and skeleton"[56][57][58][3][37] | An enigmatic herbivorous dinosauromorph.[59][3] Originally described as the earliest species of ornithischian dinosaur in 1967,[55][60][61][37] though many recent studies consider it to be a non-dinosaurian dinosauriform, potentially a silesaurid.[35][57][58][62] Under the hypothesis that ornithischians are descended from silesaurids, these different positions are not mutually exclusive.[63] |  |
| Saurosuchus[64] | S. galilei | San Juan | Cancha de Bochas,[12] lower Valle de la Luna[12] |
Hyperodapedon-Exaeretodon-Herrerasaurus biozone,[12] Exaeretodon biozone[12] |
A large number of specimens,[12][65] including two well-preserved skulls.[66][67] | A fairly common basal loricatan,[68] part of the group historicaly known as "rauisuchians". By far the largest carnivore from the formation, with a total estimated body length of around 5.5 to 7 meters (18 to 23 ft). Like most other "rauisuchians", it was a quadrupedal predator, with serrated teeth, a laterally-compressed skull, and two rows of small osteoderms running down its back.[66][67][65] |  |
| Sillosuchus[69] | S. longicervix | San Juan | middle Cancha de Bochas[12] | Hyperodapedon-Exaeretodon-Herrerasaurus biozone | A partial skeleton missing the skull,[69] and other isolated postcranial material.[12][68] | A large shuvosaurid poposauroid, the only named shuvosaurid known from outside North America.[68] The holotype individual had an estimated length of 3 m (9.8 ft),[69] but an isolated vertebra referred to this species may belong to an individual up to 10 m (33 ft) long, making it one of the largest known pseudosuchians.[68] |  |
| Trialestes | T. romeri | San Juan,[19] |
Cancha de Bochas[19] | Hyperodapedon-Exaeretodon-Herrerasaurus biozone, Teyumbaita biozone,[3] |
Two partial skeletons. The holotype includes a partial skull while a referred specimen is semi-articulated.[19] | A medium-sized crocodylomorph, one of the earliest members of the group.[19][70] Once believed to be a primitive dinosaur due to its lightly-built body plan. | 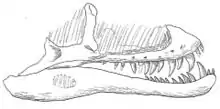 |
| Venaticosuchus | V. rusconii | La Rioja | Unknown ("middle or upper levels")[12] | Teyumbaita biozone,[3] | Partial skull with forelimb material and osteoderms.[12][71] | A medium-sized ornithosuchid, a type of scavenging[71] or piscivorous[72] pseudosuchian reaching up to 2 m (6.6 ft) in length.[71] |  |
| An unnamed lagerpetid | Unnamed | San Juan | Cancha de Bochas[12] | Hyperodapedon-Exaeretodon-Herrerasaurus biozone | Distal end of the left femur[73][12][74] | A lightly-built pterosauromorph[75][76] based on a femur discovered among the skeletal remains of the dinosaur Panphagia.[77] | |
| An unnamed crocodylomorph[70] | Unnamed | San Juan | Cancha de Bochas[70] | Hyperodapedon-Exaeretodon-Herrerasaurus biozone | Five incomplete skeletons (specimens PVSJ 846, PVSJ 1078, PVSJ 1088, PVSJ 1089 and PVSJ 1090)[70] | A new undescribed species of large-bodied crocodylomorph distinct from Trialestes.[70] | |
Other reptiles
| Non-archosaur reptiles of the Ischigualasto Formation[36][12] | |||||||
|---|---|---|---|---|---|---|---|
| Genus | Species | Province | Member | Biozone | Material | Notes | Images |
| Hyperodapedon | H. sanjuanensis[78] | San Juan, La Rioja[3] |
La Peña,[12] Cancha de Bochas[12] |
Hyperodapedon-Exaeretodon-Herrerasaurus biozone, Hyperodapedon biozone[3] |
Numerous (over 400)[12] fossils including complete skulls and postcranial bones.[3][79][80] | A large hyperodapedontine rhynchosaur. Formerly considered a species of Scaphonyx, a wastebasket taxon of large rhynchosaurs.[78] By far the most abundant species in the formation, especially in the La Peña member, though its abundance declines exponentially past the earliest layers.[14][12] Like other advanced rhynchosaurs, it was a herbivore combining a protruding beak with multi-rowed tooth plates on the side of the jaw. |  |
| H. sp. nov.[3] | La Rioja[3] | Unknown | Hyperodapedon biozone[3] | A partial skull and postcrania[3] | A new unnamed species of Hyperodapedon from the Cerro Las Lajas area.[3] More potential new species may be present due to the fact that several rhynchosaur tooth plate morphotypes are present in the formation.[81] | 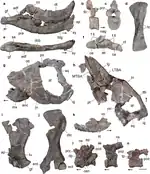 | |
| Proterochampsa | P. barrionuevoi | San Juan, La Rioja[3] |
upper La Peña,[12] |
Hyperodapedon-Exaeretodon-Herrerasaurus biozone, Teyumbaita biozone,[3] Exaeretodon biozone[12] |
Multiple skulls, some of which are complete, and associated postcranial material.[82][83][84][3] | A large and well-described proterochampsid archosauriform, with a massive flattened and ornamented skull averaging 50 centimetres (20 in) in length.[84] | 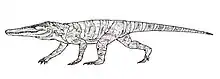 |
| Pseudochampsa[85] | P. ischigualastensis[86] | San Juan[86] | lower Cancha de Bochas[12] | Hyperodapedon-Exaeretodon-Herrerasaurus biozone | An articulated partial skeleton missing only the forelimbs.[86] | A small proterochampsid archosauriform known from a well-preserved skeleton. Originally described a species of Chanaresuchus,[86] and subsequently given its own genus.[85] |  |
| Taytalura[87] | T. alcoberi | San Juan[87] | lower Cancha de Bochas[87] | Hyperodapedon-Exaeretodon-Herrerasaurus biozone | Partial skull.[87] | A small stem-lepidosaur, distantly related to modern squamates (such as lizards) and sphenodonts (such as the tuatara).[87] | |
| Teyumbaita | T. sp.[3] | La Rioja[3] | Unknown | Teyumbaita biozone[3] | Numerous cranial and postcranial fossils, including a well-preserved partial skull.[3] | A new unnamed species of the hyperodapedontine rhynchosaur Teyumbaita, which succeeds Hyperodapedon sanjuanensis in the Cerro Las Lajas area.[3] | 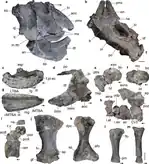 |
Synapsids
Cynodonts
| Cynodonts of the Ischigualasto Formation | |||||||
|---|---|---|---|---|---|---|---|
| Genus | Species | Province | Member | Biozone | Material | Notes | Images |
| Chiniquodon | C. sanjuanensis[88] | San Juan[88] | Cancha de Bochas[12] | Hyperodapedon-Exaeretodon-Herrerasaurus biozone | Complete skull with jaws and partial postcranial skeleton.[88] | A chiniquodontid Initially named as a species of Probelesodon,[88] a genus which is now understood to be a junior synonym of Chiniquodon.[89] | |
| C. cf. theotonicus[90] | San Juan[90] | Cancha de Bochas[12] | Hyperodapedon-Exaeretodon-Herrerasaurus biozone | Three partial skulls.[90][91][12] | A chiniquodontid, a dog-sized carnivore similar in ecological niche to some of the predatory dinosaurs it coexisted with.[92] |  | |
| Diegocanis[92] | D. elegans | San Juan[92] | Cancha de Bochas[92] | Hyperodapedon-Exaeretodon-Herrerasaurus biozone | Partial skull, represented by the snout and the orbital region, with partially preserved upper dentition.[92] | An obscure ecteniniid.[92] | |
| Ecteninion[93] | E. lunensis | San Juan[12] | Cancha de Bochas[12] | Hyperodapedon-Exaeretodon-Herrerasaurus biozone | Complete skull with jaws[93] and at least three additional specimens.[92] | A ecteniniid known from a skull around 11 centimetres (4.3 in) in length.[93] | |
| Exaeretodon | E. argentinus | San Juan, La Rioja[3] |
La Peña,[12] Cancha de Bochas,[12] Valle de la Luna[12] |
Hyperodapedon-Exaeretodon-Herrerasaurus biozone, Hyperodapedon biozone,[3] Teyumbaita biozone,[3] |
Numerous (over 200)[12] specimens, including a large number of complete skulls. | A large gomphodontosuchine traversodont, up to 1.8 meters (5.9 feet) in length. The most abundant synapsid in the Ischigualasto Formation and the second most abundant species overall, behind Hyperodapedon sanjuanensis. Like other traversodontids, it was a herbivore with a specialized chewing style. E. argentinus has a long list of potential junior synonyms,[91][12] with Exaeretodon frenguellii as a particularly widespread alternative name.[91] |  |
| cf. Probainognathus[94] | Indeterminate | San Juan[12] | Cancha de Bochas[12] | Hyperodapedon-Exaeretodon-Herrerasaurus biozone | Specimen PVSJ 410, a skull with jaws.[94][12] | A small probainognathid close to the origin of mammaliaforms. The Ischigualasto skull provisionally referred to this genus is tiny, 3.5 centimetres (1.4 in) long. It is a juvenile, presumably belonging to an unnamed species.[12] |  |
| Pseudotherium[77] | P. argentinus | San Juan[77] | upper La Peña[77] | Hyperodapedon-Exaeretodon-Herrerasaurus biozone | Specimen PVSJ 882, a cranium.[12][77][95] | A prozostrodontid probainognathian closely related to tritylodontids. A small species with a narrow skull 6.9 centimetres (2.7 in) in length, discovered among the skeletal remains of the dinosaur Panphagia.[77][95] | |
Dicynodonts
| Dicynodonts of the Ischigualasto Formation | |||||||
|---|---|---|---|---|---|---|---|
| Genus | Species | Province | Member | Biozone | Material | Notes | Images |
| Ischigualastia[96] | I. jenseni | San Juan[96] | Cancha de Bochas[12] | Hyperodapedon-Exaeretodon-Herrerasaurus biozone | Over 20 specimens,[12] five of which have been described: a complete undistorted skull, two complete compressed skulls, a partial skull with braincase material, and an ilium. Postcranial material is also present in each skull-bearing specimen.[96] | An enormous stahleckeriine stahleckeriid. The skull is 55 centimetres (22 in) in length, tall and broad, with a short snout lacking tusks. The fourth most common species in the formation. |  |
| Jachaleria | J. colorata | San Juan[12] | Quebrada de la Sal[12] | Jachaleria biozone[14][12] | Nearly complete skull with jaws.[12] | A large stahleckeriid restricted to the uppermost layers of the formation, distinct from all other Ischigualasto fossils.[14][12] Brazilian species of Jachaleria reach around 3 metres (9.8 ft) in length with an estimated mass of 300 kilograms (660 lb), close in size to Dinodontosaurus.[12] |  |
Temnospondyls
| Temnospondyls of the Ischigualasto Formation | |||||||
|---|---|---|---|---|---|---|---|
| Genus | Species | Province | Member | Biozone | Material | Notes | Images |
| Pelorocephalus[97] | P. ischigualastensis | San Juan | lower Cancha de Bochas[12] | Hyperodapedon-Exaeretodon-Herrerasaurus biozone | "A skull with associated mandibles and interclavicle."[12] | A chigutisaurid based on fragmentary material. The largest individuals are estimated to have been over 107 centimetres (42 in) in length. |  |
| Promastodonsaurus[98] | P. bellmanni | San Juan | middle Valle de la Luna[12] | Exaeretodon biozone[12] | Skull, left clavicle, interclavicle.[12] | A little-known mastodonsaurid (capitosaur). | |
Plants
Palynomorphs
Palynomorphs are often separated into two biogeographic groups in Carnian-Norian Gondwana, using Australia as a guideline for distinguishing the two.[15][16] One group is the Ipswich province, a temperate palynoflora (>40° S) which developed in areas around the Triassic South Pole, equivalent to modern South Africa, southern Australia, most of Antarctica, and the southern part of South America. The second group is the Onslow province, a subtropical palynoflora (30-40° S) which is found further north in areas closer to the Tethys Sea, equivalent to India, Madagascar, East Africa, and northern Australia.[15]
The Ischigualasto Formation, at around 40° S, is predicted to lie near the boundary between these two provinces. Palynomorphs have been found in the lower-mid part of the Valle de la Luna Member, about 330-350 meters above the base of the formation.[15][16] There are strong similarities between the Ischigualasto Formation palynoflora and the Onslow province, as well as the palynoflora of Europe. Although the most abundant pollen types are standard among Triassic South America, eight rarer palynomorph species were previously unreported from the continent.[15] Ischigualasto was the first formation reported to host distinctive Onslow province species in South America, and Western Gondwana as a whole. It helps to support the idea that the Onslow province rings around the middle latitudes of the entire Southern Hemisphere, rather than just the vicinity of the Tethys Sea.[15][16]
Onslow province species have also been discovered in the Chañares Formation, but not the Los Rastros Formation, which retains South Polar species typical of the Ipswich province.[99] This seems to contradict evidence from climate proxies which suggest the Los Rastros Formation was warmer and wetter than the Ischigualasto and Chañares formations, rather than cooler. The distinction between the Ipswich and Onslow palynofloras may be based on local environmental conditions (such as the availability of riparian or upland habitats) rather than regional climate.[5]
Macrofossils
| Plant macrofossils of the Ischigualasto Formation | ||||||
|---|---|---|---|---|---|---|
| Genus | Species | Province | Member | Material | Notes | Images |
| Agathoxylon | A. argentinum[100] | Valle de la Luna[100] | Silica permineralizations of wood.[100] | An araucariacean conifer. | ||
| Araucarioxylon | A. sp. A | An araucarian.[27] | ||||
| Cladophlebis | C. mendozaensis | A fern.[27] | ||||
| Dicroidium | D. lancifolium | Horizontally oriented cuticles and impressions, as well as cuticles oriented obliquely with respect to the bedding plane.[27] | A seed fern.[27] | |||
| D. odontopteroides | Horizontally oriented cuticles and impressions, as well as cuticles oriented obliquely with respect to the bedding plane.[27] | A seed fern.[27] |  | |||
| D. zuberi[101] | San Juan | Valle de la Luna | Isolated stacks of leaf cuticle, so-called 'litter bags'.[101] | A seed fern,[101] previously described as a species of Zuberia.[27] | ||
| Ginkgoites | G. sp. | A ginkgoale.[27] |  | |||
| Heidiphyllum | H. elongatum | A podocarp.[27] | ||||
| Johnstonia | J. coriacea | A seed fern.[27] | ||||
| J. stelzneriana | Horizontally oriented cuticles and impressions, as well as cuticles oriented obliquely with respect to the bedding plane.[27] | A seed fern.[27] | ||||
| Lepidopteris | L. stormbergensis | A seed fern.[27] | ||||
| Michelilloa | M. waltonii | A cycad.[27][102] | ||||
| Neocalamites | N. carrerei | Horizontally oriented cuticles and impressions.[27] | A horsetail.[27] | |||
| Protojuniperoxylon | P. ischigualastensis | A conifer.[27] | ||||
| Pterophyllum | P. sp. | A bennettitale.[27] |  | |||
| Rhexoxylon | R. piatnitzkyi | Vertical silicified trunks and stumps as well as horizontal or oblique silicified tree trunks.[27] | A seed fern.[27][103] | |||
| Scytophyllum | S. neubergianum | A seed fern.[27] | ||||
| Taeniopteris | T. sp. | A possible cycadophyte.[27] | ||||
| Xylopteris | X. argentina | Horizontally oriented cuticles and impressions, as well as cuticles oriented obliquely with respect to the bedding plane.[27] | A seed fern.[27] | |||
| X. elongata | Horizontally oriented cuticles and impressions, as well as cuticles oriented obliquely with respect to the bedding plane.[27] | A seed fern.[27] | ||||
| Yabiella | Y. brackebuschiana | A cycad.[27] | ||||
| Y. mareyesiaca | A cycad.[27] | |||||
| Y. sp. | A cycad.[27] | |||||
| Zuberia | Z. papillata | A corystosperm seed fern similar to Dicroidium.[104][27] | ||||
See also
- Quebrada del Barro Formation, contemporaneous fossiliferous formation of the Marrayel-El Carrizal Basin
- Candelária Formation, contemporaneous fossiliferous formation of the Paraná Basin
- Molteno Formation, contemporaneous fossiliferous formation of the Karoo Basin in southern Africa
- Fremouw Formation, contemporaneous fossiliferous formation of Antarctica
- Denmark Hill Insect Bed, contemporaneous fossiliferous unit of Queensland, Australia
- Madygen Formation, contemporaneous Lagerstätte of Central Asia
References
- ↑ (in Spanish) El lugar donde se posa la luna Archived 2019-03-29 at the Wayback Machine
- 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 Currie, Brian S.; Colombi, Carina E.; Tabor, Neil J.; Shipman, Todd C.; Montañez, Isabel P. (2009). "Stratigraphy and architecture of the Upper Triassic Ischigualasto Formation, Ischigualasto Provincial Park, San Juan, Argentina". Journal of South American Earth Sciences. 27 (1): 74–87. doi:10.1016/j.jsames.2008.10.004.
- 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 Julia B. Desojo; Lucas E. Fiorelli; Martín D. Ezcurra; Agustín G. Martinelli; Jahandar Ramezani; Átila. A. S. Da Rosa; M. Belén von Baczko; M. Jimena Trotteyn; Felipe C. Montefeltro; Miguel Ezpeleta; Max C. Langer (2020). "The Late Triassic Ischigualasto Formation at Cerro Las Lajas (La Rioja, Argentina): fossil tetrapods, high-resolution chronostratigraphy, and faunal correlations". Scientific Reports. 10 (1): Article number 12782. Bibcode:2020NatSR..1012782D. doi:10.1038/s41598-020-67854-1. PMC 7391656. PMID 32728077.
- ↑ Balabusic et al., 2001, p.26
- 1 2 3 4 5 6 7 Mancuso, Adriana C.; Irmis, Randall B.; Pedernera, Tomás E.; Gaetano, Leandro C.; Benavente, Cecilia A.; Breeden III, Benjamin T. (2022-06-13). "Paleoenvironmental and Biotic Changes in the Late Triassic of Argentina: Testing Hypotheses of Abiotic Forcing at the Basin Scale". Frontiers in Earth Science. 10. doi:10.3389/feart.2022.883788. ISSN 2296-6463.
- ↑ Schencman, 2015, p.220
- ↑ Aceituno Cieri et al., 2015, p.60
- ↑ Monetta et al., 2000, p.644
- ↑ Colombi, Carina E.; Limarino, Carlos O.; Alcober, Oscar A. (2017). "Allogenic controls on the fluvial architecture and fossil preservation of the Upper Triassic Ischigualasto Formation, NW Argentina". Sedimentary Geology. 362: 1–16. doi:10.1016/j.sedgeo.2017.10.003.
- 1 2 3 Rogers, Raymond R.; Swisher, Carl C.; Sereno, Paul C.; Monetta, Alfredo M.; Forster, Catherine A.; Martínez, Ricardo N. (1993). "The Ischigualasto Tetrapod Assemblage (Late Triassic, Argentina) and 40Ar/39Ar Dating of Dinosaur Origins". Science. 260 (5109): 794–797. doi:10.1126/science.260.5109.794. ISSN 0036-8075. PMID 17746113. S2CID 35644127.
- 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 Tabor, Neil J.; Montañez, Isabel P.; Kelso, Kelley A.; Currie, Brian; Shipman, Todd; Colombi, Carina (2006), "A Late Triassic soil catena: Landscape and climate controls on paleosol morphology and chemistry across the Carnian-age Ischigualasto–Villa Union basin, northwestern Argentina", Paleoenvironmental Record and Applications of Calcretes and Palustrine Carbonates, Geological Society of America, doi:10.1130/2006.2416(02), ISBN 978-0-8137-2416-4
- 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 63 64 65 66 67 68 69 70 Martínez, Ricardo N.; Apaldetti, Cecilia; Alcober, Oscar A.; Colombi, Carina E.; Sereno, Paul C.; Fernandez, Eliana; Malnis, Paula Santi; Correa, Gustavo A.; Abelin, Diego (2013). "Vertebrate succession in the Ischigualasto Formation". Journal of Vertebrate Paleontology. 32 (Memoir 12: Basal sauropodomorphs and the vertebrate fossil record of the Ischigualasto Formation (Late Triassic: Carnian-Norian) of Argentina, sup1): 10–30. doi:10.1080/02724634.2013.818546. hdl:11336/7771. ISSN 0272-4634. S2CID 37918101.
- ↑ Wallace, 2018, p.6
- 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 Martinez, Ricardo N.; Sereno, Paul C.; Alcober, Oscar A.; Colombi, Carina E.; Renne, Paul R.; Montañez, Isabel P.; Currie, Brian S. (2011). "A Basal Dinosaur from the Dawn of the Dinosaur Era in Southwestern Pangaea". Science. 331 (6014): 206–210. doi:10.1126/science.1198467. hdl:11336/69202. ISSN 0036-8075. PMID 21233386. S2CID 33506648.
- 1 2 3 4 5 6 7 Césari, Silvia N.; Colombi, Carina E. (2013-05-21). "A new Late Triasssic phytogeographical scenario in westernmost Gondwana". Nature Communications. 4 (1): 1889. doi:10.1038/ncomms2917. ISSN 2041-1723. PMID 23695683. S2CID 205317325.
- 1 2 3 4 5 Césari, Silvia N.; Colombi, Carina (2016). "Palynology of the Late Triassic Ischigualasto Formation, Argentina: Paleoecological and paleogeographic implications". Palaeogeography, Palaeoclimatology, Palaeoecology. 449: 365–384. doi:10.1016/j.palaeo.2016.02.023.
- 1 2 Irmis, Randall B.; Mundil, Roland; Mancuso, Adriana Cecilia; Carrillo-Briceño, Jorge D.; Ottone, Eduardo G.; Marsicano, Claudia A. (2022). "South American Triassic geochronology: Constraints and uncertainties for the tempo of Gondwanan non-marine vertebrate evolution". Journal of South American Earth Sciences. 116: 103770. doi:10.1016/j.jsames.2022.103770.
- 1 2 Colombi, Carina; Martínez, Ricardo N.; Césari, Silvia N.; Alcober, Oscar; Limarino, Carlos O.; Montañez, Isabel (2021-11-01). "A high-precision U–Pb zircon age constraints the timing of the faunistic and palynofloristic events of the Carnian Ischigualasto Formation, San Juan, Argentina". Journal of South American Earth Sciences. 111: 103433. doi:10.1016/j.jsames.2021.103433. ISSN 0895-9811.
- 1 2 3 4 5 6 Lecuona, Agustina; Ezcurra, Martín D.; Irmis, Randall B. (2016). "Revision of the early crocodylomorph Trialestes romeri (Archosauria, Suchia) from the lower Upper Triassic Ischigualasto Formation of Argentina: one of the oldest‐known crocodylomorphs". Papers in Palaeontology. 2 (4): 585–622. doi:10.1002/spp2.1056. ISSN 2056-2799. S2CID 88719030.
- ↑ Fernanda, Oliveira da Silva; Martinelli, Agustín G.; Ferigolo, Jorge; Ribeiro, Ana Maria (2023), "A new herrerasaurid dinosaur record from southern Brazil (Upper Triassic) and its faunal association", Revista Brasileira de Paleontologia, 26 (2): 97–113, doi:10.4072/rbp.2023.2.02, S2CID 259942523
- ↑ Mancuso, Adriana C.; Benavente, Cecilia A.; Irmis, Randall B.; Mundil, Roland (2020). "Evidence for the Carnian Pluvial Episode in Gondwana: New multiproxy climate records and their bearing on early dinosaur diversification". Gondwana Research. 86: 104–125. doi:10.1016/j.gr.2020.05.009. ISSN 1342-937X.
- ↑ Benavente, C.A.; Mancuso, A.C.; Irmis, R.B.; Bohacs, K.M.; Matheos, S. (2021-03-29). "Tectonically conditioned record of continental interior paleoclimate during the Carnian Pluvial Episode: The Upper Triassic Los Rastros Formation, Argentina". GSA Bulletin. 134 (1–2): 60–80. doi:10.1130/b35847.1. ISSN 0016-7606.
- ↑ Colombi, Carina; Martínez, Ricardo N.; Césari, Silvia N.; Alcober, Oscar; Limarino, Carlos O.; Montañez, Isabel (2021-11-01). "A high-precision U–Pb zircon age constraints the timing of the faunistic and palynofloristic events of the Carnian Ischigualasto Formation, San Juan, Argentina". Journal of South American Earth Sciences. 111: 103433. doi:10.1016/j.jsames.2021.103433. ISSN 0895-9811.
- 1 2 Mancuso, Adriana Cecilia; Horn, Bruno Ludovico Dihl; Benavente, Cecilia Andrea; Schultz, Cesar Leandro; Irmis, Randall B. (2021). "The paleoclimatic context for South American Triassic vertebrate evolution". Journal of South American Earth Sciences. 110: 103321. doi:10.1016/j.jsames.2021.103321.
- ↑ Mancuso, Adriana C.; Irmis, Randall B.; Pedernera, Tomás E.; Gaetano, Leandro C.; Benavente, Cecilia A.; Breeden III, Benjamin T. (2022-06-13). "Paleoenvironmental and Biotic Changes in the Late Triassic of Argentina: Testing Hypotheses of Abiotic Forcing at the Basin Scale". Frontiers in Earth Science. 10. doi:10.3389/feart.2022.883788. ISSN 2296-6463.
{{cite journal}}: CS1 maint: unflagged free DOI (link) - 1 2 3 Tabor, Neil J.; Montañez, Isabel P.; Zierenberg, Robert; Currie, Brian S. (2004). "Mineralogical and geochemical evolution of a basalt-hosted fossil soil (Late Triassic, Ischigualasto Formation, northwest Argentina): Potential for paleoenvironmental reconstruction". Geological Society of America Bulletin. 116 (9): 1280. doi:10.1130/B25222.1. ISSN 0016-7606.
- 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 Colombi, Carina E.; Parrish, Judith Totman (2008). "Late Triassic Environmental Evolution in Southwestern Pangea: Plant Taphonomy of the Ischigualasto Formation". PALAIOS. 23 (12): 778–795. Bibcode:2008Palai..23..778C. doi:10.2110/palo.2007.p07-101r. S2CID 129121885.
- ↑ Colombi, Carina E.; Parrish, Judith Totman (2008). "Late Triassic Environmental Evolution in Southwestern Pangea: Plant Taphonomy of the Ischigualasto Formation". PALAIOS. 23 (12): 778–795. Bibcode:2008Palai..23..778C. doi:10.2110/palo.2007.p07-101r. S2CID 129121885.
{{cite journal}}: CS1 maint: url-status (link) - ↑ Mancuso, Adriana Cecilia; Previtera, Elena; Benavente, Cecilia Andrea; Del Pino, Santiago Hernandez (2017). "EVIDENCE OF BACTERIAL DECAY AND EARLY DIAGENESIS IN A PARTIALLY ARTICULATED TETRAPOD FROM THE TRIASSIC CHAÑARES FORMATION". PALAIOS. 32 (6): 367–381. doi:10.2110/palo.2016.076. hdl:11336/63822. ISSN 0883-1351. S2CID 134067728.
- ↑ Sagasti, Ana Julia; Bodnar, Josefina (2023). "Biological decay by microorganisms in stems from the Upper Triassic Ischigualasto Formation (San Juan Province, Argentina): A striking microbial diversity in Carnian-Norian terrestrial ecosystems". Review of Palaeobotany and Palynology. 315: 104915. doi:10.1016/j.revpalbo.2023.104915. ISSN 0034-6667. S2CID 258779705.
- ↑ Colombi, Carina E.; Rogers, Raymond R.; Alcober, Oscar A. (2012). "Vertebrate taphonomy of the Ischigualasto Formation" (PDF). Journal of Vertebrate Paleontology. 32 (sup1): 31–50. doi:10.1080/02724634.2013.809285. ISSN 0272-4634. S2CID 131586488.
- ↑ Hollocher, K. T.; Alcober, O. A.; Colombi, C. E.; Hollocher, T. C. (2005). "Carnivore Coprolites from the Upper Triassic Ischigualasto Formation, Argentina: Chemistry, Mineralogy, and Evidence for Rapid Initial Mineralization". PALAIOS. 20 (1): 51–63. doi:10.2110/palo.2003.p03-98. ISSN 0883-1351. S2CID 131242248.
- 1 2 Colombi, Carina E.; Jofré, Carolina; Currie, Brian S. (2008). "Large-diameter burrows in the Upper Triassic Ischigualasto Formation, Northwestern Argentina". Ameghiniana. 45 (4): 795–799.
- 1 2 3 Colombi, Carina E.; Fernández, Eliana; Currie, Brian S.; Alcober, Oscar A.; Martínez, Ricardo; Correa, Gustavo (2012-12-05). "Large-Diameter Burrows of the Triassic Ischigualasto Basin, NW Argentina: Paleoecological and Paleoenvironmental Implications". PLOS ONE. 7 (12): e50662. doi:10.1371/journal.pone.0050662. ISSN 1932-6203. PMC 3515583. PMID 23227195.
- 1 2 3 4 5 Weishampel, Dodson & Osmólska 2004, pp. 527–528.
- 1 2 3 4 5 6 7 8 9 10 11 12 Ezcurra, Martin D. (2010). "A new early dinosaur (Saurischia: Sauropodomorpha) from the Late Triassic of Argentina: a reassessment of dinosaur origin and phylogeny". Journal of Systematic Palaeontology. 8 (3): 371–425. doi:10.1080/14772019.2010.484650. ISSN 1477-2019. S2CID 129244872.
- 1 2 3 4 5 6 7 8 9 10 Novas, Fernando E.; Agnolin, Federico L.; Ezcurra, Martín D.; Temp Müller, Rodrigo; Martinelli, Agustín G.; Langer, Max C. (2021-10-01). "Review of the fossil record of early dinosaurs from South America, and its phylogenetic implications". Journal of South American Earth Sciences. 110: 103341. doi:10.1016/j.jsames.2021.103341. ISSN 0895-9811.
- 1 2 3 Sereno, Paul C.; Forster, Catherine A.; Rogers, Raymond R.; Monetta, Alfredo M. (1993). "Primitive dinosaur skeleton from Argentina and the early evolution of Dinosauria". Nature. 361 (6407): 64–66. doi:10.1038/361064a0. ISSN 0028-0836. S2CID 4270484.
- 1 2 3 4 Sereno, Paul C.; Martínez, Ricardo N.; Alcober, Oscar A. (2013). "Osteology of Eoraptor lunensis (Dinosauria, Sauropodomorpha). Basal sauropodomorphs and the vertebrate fossil record of the Ischigualasto Formation (Late Triassic: Carnian-Norian) of Argentina". Journal of Vertebrate Paleontology Memoir. 12: 83–179. doi:10.1080/02724634.2013.820113. JSTOR 24413073. S2CID 86006363.
- 1 2 Weishampel, Dodson & Osmólska 2004, p. 26, Table 2.1.
- 1 2 Sereno, P.C.; Novas, F.E. (1992). "The complete skull and skeleton of an early dinosaur". Science. 258 (5085): 1137–1140. Bibcode:1992Sci...258.1137S. doi:10.1126/science.258.5085.1137. PMID 17789086. S2CID 1640394.
- ↑ Reig, O.A. (1963). "La presencia de dinosaurios saurisquios en los "Estratos de Ischigualasto" (Mesotriásico Superior) de las provincias de San Juan y La Rioja (República Argentina)". Ameghiniana (in Spanish). 3 (1): 3–20.
- ↑ Sereno, P.C. (1993). "The pectoral girdle and forelimb of the basal theropod Herrerasaurus ischigualastensis". Journal of Vertebrate Paleontology. 13 (4): 425–450. doi:10.1080/02724634.1994.10011524.
- ↑ Novas, F.E. (1993). "New information on the systematics and postcranial skeleton of Herrerasaurus ischigualastensis (Theropoda: Herrerasauridae) from the Ischigualasto Formation (Upper Triassic) of Argentina". Journal of Vertebrate Paleontology. 13 (4): 400–423. doi:10.1080/02724634.1994.10011523.
- ↑ Sereno, P.C.; Novas, F.E. (1993). "The skull and neck of the basal theropod Herrerasaurus ischigualastensis". Journal of Vertebrate Paleontology. 13 (4): 451–476. doi:10.1080/02724634.1994.10011525.
- 1 2 3 4 5 Martinez, Ricardo N.; Alcober, Oscar A. (2009). Sereno, Paul (ed.). "A Basal Sauropodomorph (Dinosauria: Saurischia) from the Ischigualasto Formation (Triassic, Carnian) and the Early Evolution of Sauropodomorpha". PLOS ONE. 4 (2): e4397. doi:10.1371/journal.pone.0004397. ISSN 1932-6203. PMC 2635939. PMID 19209223.
- ↑ Cabreira, Sergio F.; Schultz, Cesar L.; Bittencourt, Jonathas S.; Soares, Marina B.; Fortier, Daniel C.; Silva, Lúcio R.; Langer, Max C. (2011). "New stem-sauropodomorph (Dinosauria, Saurischia) from the Triassic of Brazil" (PDF). Naturwissenschaften. 98 (12): 1035–1040. doi:10.1007/s00114-011-0858-0. ISSN 0028-1042. PMID 22083251. S2CID 5721100.
- 1 2 3 4 5 Martínez, Ricardo; Alcober, Oscar (2010). "A new herrerasaurid (Dinosauria, Saurischia) from the Upper Triassic Ischigualasto Formation of northwestern Argentina". ZooKeys (63): 55–81. doi:10.3897/zookeys.63.550. ISSN 1313-2970. PMC 3088398. PMID 21594020.
- 1 2 3 4 5 6 Desojo, J.B.; Ezcurra, M.D. (2011). "A reappraisal of the taxonomic status of Aetosauroides (Archosauria, Aetosauria) specimens from the Late Triassic of South America and their proposed synonymy with Stagonolepis". Journal of Vertebrate Paleontology. 31 (3): 596–609. doi:10.1080/02724634.2011.572936. S2CID 85418778.
- ↑ Tabora, J.R.A.; Heckert, A.B.; Desojo, J.B. (2015). "Intraspecific variation in Aetosauroides Casamiquela (Archosauria: Aetosauria) from the Upper Triassic of Argentina and Brazil: an example of sexual dimorphism?". Ameghiniana. 52 (2): 173–187. doi:10.5710/AMGH.05.01.2015.2824. S2CID 11498039.
- ↑ Lucas, S.G.; Heckert, A.B. (1996). "Late Triassic aetosaur biochronology" (PDF). Albertiana. 17: 57–64. Archived from the original (PDF) on 2013-11-13. Retrieved 2011-05-11.
- ↑ Heckert, A.B.; Lucas, S.G. (2002). "South American occurrences of the Adamanian (Late Triassic: latest Carnian) index taxon Stagonolepis (Archosauria: Aetosauria) and their biochronological significance". Journal of Paleontology. 76 (5): 852–863. doi:10.1666/0022-3360(2002)076<0852:SAOOTA>2.0.CO;2. S2CID 128610620.
- 1 2 Heckert, Andrew B.; Martínez, Ricardo N.; Celeskey, Matthew D. (2021-10-31). "Anatomical Details of Aetosauria (Archosauria: Pseudosuchia) as Revealed by an Articulated Posterior Skeleton from the Upper Triassic Ischigualasto Formation, San Juan Province, Argentina". Ameghiniana. 58 (6): 464–484. doi:10.5710/AMGH.05.09.2021.3426. ISSN 0002-7014. S2CID 239751891.
- 1 2 3 4 Imanol Yáñez; Diego Pol; Juan Martín Leardi; Oscar A. Alcober; Ricardo N. Martínez (2021). "An enigmatic new archosauriform from the Carnian– Norian, Upper Triassic, Ischigualasto Formation of northwestern Argentina". Acta Palaeontologica Polonica. 66 (3): 509–533. doi:10.4202/app.00806.2020.
- 1 2 Casamiquela, R.M. (1967). "Un nuevo dinosaurio ornitisquio triásico (Pisanosaurus mertii; Ornithopoda) de la Formación Ischigualasto, Argentina". Ameghiniana. 4 (2): 47–64.
- ↑ Weishampel, Dodson & Osmólska 2004, p. 326, Table 14.1.
- 1 2 Federico L. Agnolin (2015). "Nuevas observaciones sobre Pisanosaurus mertii Casamiquela, 1967 (Dinosauriformes) y sus implicancias taxonómicas" (PDF). XXIX Jornadas Argentinas de Paleontología de Vertebrados. 27–29 de Mayo de 2015. Diamante, Entre Ríos. Libro de Resúmenes: 13–14. Archived from the original (PDF) on 2016-01-27. Retrieved 2015-08-07.
- 1 2 Agnolín, Federico L.; Rozadilla, Sebastián (2017). "Phylogenetic reassessment of Pisanosaurus mertii Casamiquela, 1967, a basal dinosauriform from the Late Triassic of Argentina". Journal of Systematic Palaeontology. 16 (10): 853–879. doi:10.1080/14772019.2017.1352623. S2CID 90655527.
- ↑ Matthew G. Baron (2018). "Pisanosaurus mertii and the Triassic ornithischian crisis: could phylogeny offer a solution?". Historical Biology: An International Journal of Paleobiology. 31 (8): 1–15. doi:10.1080/08912963.2017.1410705. S2CID 89924902.
- ↑ Bonaparte, J.F. (1976). "Pisanosaurus mertii Casamiquela and the origin of the Ornithischia". Journal of Paleontology. 50 (5): 808–820. JSTOR 1303575.
- ↑ Baron, M.G.; Norman, D.B.; Barrett, P.M. (2017). "A new hypothesis of dinosaur relationships and early dinosaur evolution". Nature. 543 (7646): 501–506. Bibcode:2017Natur.543..501B. doi:10.1038/nature21700. PMID 28332513. S2CID 187506290.
- ↑ Matthew G. Baron; David B. Norman; Paul M. Barrett (2017). "Baron et al. reply". Nature. 551 (7678): E4–E5. Bibcode:2017Natur.551E...4B. doi:10.1038/nature24012. PMID 29094705. S2CID 205260360.
- ↑ Müller, Rodrigo Temp; Garcia, Maurício Silva (August 2020). "A paraphyletic 'Silesauridae' as an alternative hypothesis for the initial radiation of ornithischian dinosaurs". Biology Letters. 16 (8): 20200417. doi:10.1098/rsbl.2020.0417. ISSN 1744-9561. PMC 7480155. PMID 32842895.
- ↑ Reig, O. A. (1959). "Primeros datos descriptivos sobre nuevos arcosaurios del Triásico de Ischigualasto (San Juan, Argentina)". Revista de la Asociación Geológica Argentina. 13 (4): 257–270.
- 1 2 Trotteyn, M.J.; Desojo, J.; Alcober, O. (2011). "Nuevo material postcraneano de Saurosuchus galilei (Archosauria: Crurotarsi) del Triásico Superior del centro-oeste de Argentina". Ameghiniana (in Spanish). 48 (1): 13–27. doi:10.5710/amgh.v48i1(265). hdl:11336/98158. S2CID 129032513.
- 1 2 Sill, W. D. (1974). "The anatomy of Saurosuchus galilei and the relationships of the rauisuchid thecodonts". Bulletin of the Museum of Comparative Zoology. 146: 317–362.
- 1 2 Alcober, O. (2000). "Redescription of the skull of Saurosuchus galilei (Archosauria: Rauisuchidae)". Journal of Vertebrate Paleontology. 20 (2): 302–316. doi:10.1671/0272-4634(2000)020[0302:ROTSOS]2.0.CO;2. S2CID 85967965.
- 1 2 3 4 Sterling J. Nesbitt (2011). "The Early Evolution of Archosaurs: Relationships and the Origin of Major Clades". Bulletin of the American Museum of Natural History. 352: 1–292. doi:10.1206/352.1. hdl:2246/6112. S2CID 83493714.
- 1 2 3 Alcober, Oscar; Parrish, J. Michael (1997). "A new poposaurid from the Upper Triassic of Argentina". Journal of Vertebrate Paleontology. 17 (3): 548–556. doi:10.1080/02724634.1997.10011001. ISSN 0272-4634.
- 1 2 3 4 5 Juan Martín Leardi; Imanol Yáñez; Diego Pol (2020). "South American Crocodylomorphs (Archosauria; Crocodylomorpha): A review of the early fossil record in the continent and its relevance on understanding the origins of the clade". Journal of South American Earth Sciences. 104: Article 102780. Bibcode:2020JSAES.10402780L. doi:10.1016/j.jsames.2020.102780. S2CID 225237455.
- 1 2 3 Von Baczko, M. Belén (2018). "Rediscovered Cranial Material of Venaticosuchus rusconii Enables the First Jaw Biomechanics in Ornithosuchidae (Archosauria: Pseudosuchia)". Ameghiniana. 55 (4): 365–379. doi:10.5710/AMGH.19.03.2018.3170. hdl:11336/99976. S2CID 134536703.
- ↑ Taborda, Jeremias; von Baczko, M. Belén; Desojo, Julia (2023). "Biomechanical analysis and new trophic hypothesis for Riojasuchus tenuisceps, a bizarre-snouted Late Triassic pseudosuchian from Argentina" (PDF). Acta Palaeontologica Polonica. 68. doi:10.4202/app.01038.2022. ISSN 0567-7920. S2CID 261551160.
- ↑ Langer, Max C.; Nesbitt, Sterling J.; Bittencourt, Jonathas S.; Irmis, Randall B. (2013). "Non-dinosaurian Dinosauromorpha". Geological Society, London, Special Publications. 379 (1): 157–186. doi:10.1144/sp379.9. ISSN 0305-8719.
- ↑ Müller, Rodrigo Temp; Langer, Max Cardoso; Dias-da-Silva, Sérgio (2018-03-07). "Ingroup relationships of Lagerpetidae (Avemetatarsalia: Dinosauromorpha): a further phylogenetic investigation on the understanding of dinosaur relatives". Zootaxa. 4392 (1): 149–158. doi:10.11646/zootaxa.4392.1.7. ISSN 1175-5334. PMID 29690420.
- ↑ Kammerer, Christian F.; Nesbitt, Sterling J.; Flynn, John J.; Ranivoharimanana, Lovasoa; Wyss, André R. (2020-07-02). "A tiny ornithodiran archosaur from the Triassic of Madagascar and the role of miniaturization in dinosaur and pterosaur ancestry". Proceedings of the National Academy of Sciences. 117 (30): 17932–17936. doi:10.1073/pnas.1916631117. ISSN 0027-8424. PMC 7395432. PMID 32631980.
- ↑ Ezcurra, Martín D.; Nesbitt, Sterling J.; Bronzati, Mario; Dalla Vecchia, Fabio Marco; Agnolin, Federico L.; Benson, Roger B. J.; Brissón Egli, Federico; Cabreira, Sergio F.; Evers, Serjoscha W.; Gentil, Adriel R.; et al. (2020-12-09). "Enigmatic dinosaur precursors bridge the gap to the origin of Pterosauria". Nature. 588 (7838): 445–449. doi:10.1038/s41586-020-3011-4. ISSN 0028-0836. PMID 33299179. S2CID 228077525.
- 1 2 3 4 5 6 Wallace, Rachel V. S.; Martínez, Ricardo; Rowe, Timothy (2019). "First record of a basal mammaliamorph from the early Late Triassic Ischigualasto Formation of Argentina". PLOS ONE. 14 (8): e0218791. doi:10.1371/journal.pone.0218791. ISSN 1932-6203. PMC 6685608. PMID 31390368.
- 1 2 Sill, William D. (1970). "Schaphonyx sanjuanensis, nuevo rincosaurio (Reptilia) de la Formación Ischigualasto, Triásico de San Juan Argentina". Ameghiniana (in Spanish). 7 (4): 341–354. ISSN 1851-8044.
- ↑ Gentil, Adriel R.; Ezcurra, Martín D. (2018). "Reconstruction of the Masticatory Apparatus of the Holotype of the Rhynchosaur Hyperodapedon sanjuanensis from the Late Triassic of Argentina: Implications for the Diagnosis of the Species". Ameghiniana. 55 (2): 137–149. doi:10.5710/AMGH.17.10.2017.3132. ISSN 0002-7014. S2CID 134557963.
- ↑ Gentil, Adriel R.; Ezcurra, Martín D. (2022). "Skull osteology of the holotype of the rhynchosaur Hyperodapedon sanjuanensis (Sill, 1970) from the Upper Triassic Ischigualasto Formation of Argentina". The Anatomical Record. 305 (5): 1168–1200. doi:10.1002/ar.24771. ISSN 1932-8486. PMID 34496139. S2CID 237454269.
- ↑ Gentil, Adriel R.; Ezcurra, Martín D. (2018-02-16). "A new rhynchosaur maxillary tooth plate morphotype expands the disparity of the group in the Ischigualasto Formation (Late Triassic) of Northwestern Argentina". Historical Biology: 1–8. doi:10.1080/08912963.2018.1438425. ISSN 0891-2963. S2CID 90161690.
- 1 2 Trotteyn, María Jimena; Haro, José Augusto (2011-03-01). "The braincase of a specimen of Proterochampsa Reig (Archosauriformes: Proterochampsidae) from the Late Triassic of Argentina". Paläontologische Zeitschrift. 85 (1): 1–17. doi:10.1007/s12542-010-0068-7. ISSN 1867-6812. S2CID 129156275.
- 1 2 Trotteyn, M. Jimena (2011). "Material Postcraneano de Proterochampsa barrionuevoi Reig 1959 (Diapsida: Archosauriformes) del Triásico Superior del Centro-oeste de Argentina". Ameghiniana (in Spanish). 48 (4): 424–446. doi:10.5710/AMGH.v48i4(351). hdl:11336/197236. S2CID 130001247.
- 1 2 3 Dilkes, David; Arcucci, Andrea (19 June 2012). "Proterochampsa barrionuevoi (Archosauriformes: Proterochampsia) from the Late Triassic (Carnian) of Argentina and a phylogenetic analysis of Proterochampsia". Palaeontology. 55 (4): 853–885. doi:10.1111/j.1475-4983.2012.01170.x. S2CID 129670473.
- 1 2 Trotteyn, M. Jimena; Ezcurra, Martín D. (2014). "Osteology of Pseudochampsa ischigualastensis gen. et comb. nov. (Archosauriformes: Proterochampsidae) from the Early Late Triassic Ischigualasto Formation of Northwestern Argentina". PLOS ONE. 9 (11): e111388. doi:10.1371/journal.pone.0111388. ISSN 1932-6203. PMC 4245112. PMID 25426846.
- 1 2 3 4 Trotteyn, María J.; Martínez, Ricardo N.; Alcober, Oscar A. (2012-02-28). "A new proterochampsid Chanaresuchus ischigualastensis (Diapsida, Archosauriformes) in the early Late Triassic Ischigualasto Formation, Argentina". Journal of Vertebrate Paleontology. 32 (2): 485–489. doi:10.1080/02724634.2012.645975. hdl:11336/199496. ISSN 0272-4634. S2CID 85962463.
- 1 2 3 4 5 Ricardo N. Martínez; Tiago R. Simões; Gabriela Sobral; Sebastián Apesteguía (2021). "A Triassic stem lepidosaur illuminates the origin of lizard-like reptiles". Nature. 597 (7875): 235–238. Bibcode:2021Natur.597..235M. doi:10.1038/s41586-021-03834-3. PMID 34433961. S2CID 237307957.
- 1 2 3 4 Martinez, Ricardo N.; Forster, Catherine A. (1996). "The skull of Probelesodon sanjuanensis, sp. nov., from the Late Triassic Ischigualasto Formation of Argentina". Journal of Vertebrate Paleontology. 16 (2): 285–291. doi:10.1080/02724634.1996.10011315. ISSN 0272-4634.
- ↑ Abdala, Fernando; Giannini, Norberto P. (2002). "Chiniquodontid cynodonts: systematic and morphometric considerations". Palaeontology. 45 (6): 1151–1170. doi:10.1111/1475-4983.00280. ISSN 0031-0239. S2CID 128422947.
- 1 2 3 Bonaparte, J. F. (1966-12-06). "Chiniquodon Huene (Therapsida, Cynodontia) en el Triásico de Ischigualasto, Argentina: Consideraciones sobre su asignación familiar". Acta Geológica Lilloana (in Spanish). 8: 157–169. ISSN 1852-6217.
- 1 2 3 Abdala, Fernando (2000). "Catalogue of non-mammalian cynodonts in the Vertebrate Paleontology Collection of the Instituto Miguel Lillo, Universidad Nacional de Tucumán, with comments on species". Ameghiniana. 37 (4): 463–475.
- 1 2 3 4 5 6 7 N. Martínez, Ricardo; Fernandez, Eliana; A. Alcober, Oscar (2013). "A new non-mammaliaform eucynodont from the Carnian-Norian Ischigualasto Formation, Northwestern Argentina". Revista Brasileira de Paleontologia. 16 (1): 61–76. doi:10.4072/rbp.2013.1.05. ISSN 1519-7530.
- 1 2 3 Martinez, Ricardo N.; May, Cathleen L.; Forster, Catherine A. (1996-06-05). "A new carnivorous cynodont from the Ischigualasto Formation (Late Triassic, Argentina), with comments on eucynodont phylogeny". Journal of Vertebrate Paleontology. 16 (2): 271–284. doi:10.1080/02724634.1996.10011314. ISSN 0272-4634.
- 1 2 Bonaparte, J.F.; Crompton, A.W. (1994). "A juvenile probainognathid cynodont skull from the Ischigualasto Formation and the origin of mammals". Revista del Museo Argentino Ciencias Naturales. 5 (1–2).
- 1 2 Wallace, 2018, p.10
- 1 2 3 Cox, C.B. (1965-03-04). "New Triassic dicynodonts from South America, their origins and relationships". Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. 248 (753): 457–514. doi:10.1098/rstb.1965.0005. ISSN 2054-0280. S2CID 86293260.
- ↑ Bonaparte, J.F. (1975). "Sobre la presencia del laberintodonte Pelorocephalus en la Formacion de Ischigualasto y su significado estratigrafico". Adas 1 Congress Argentinoda Paleontologia y Biostratigrafica (in Spanish). Tucuman: Asociacion Paleontologica Argentina. pp. 537–544.
- ↑ Bonaparte, José F. (1963). "Promastodonsaurus bellmani, capitosáurido del Triásico medio de Argentina". Ameghiniana (in Spanish). 3 (3): 67–78.
- ↑ Perez Loinaze, Valeria S.; Vera, Ezequiel I.; Fiorelli, Lucas E.; Desojo, Julia B. (2018). "Palaeobotany and palynology of coprolites from the Late Triassic Chañares Formation of Argentina: implications for vegetation provinces and the diet of dicynodonts". Palaeogeography, Palaeoclimatology, Palaeoecology. 502: 31–51. doi:10.1016/j.palaeo.2018.04.003. S2CID 134075049.
- 1 2 3 Bodnar, Josefina; Sagasti, Ana Julia; Correa, Gustavo A.; Miranda, Victoria; Medina, Florencia (2022). "Araucariaceous fossil woods from the Upper Triassic Ischigualasto Formation (San Juan Province, Argentina): paleofloristic and paleoclimatic implications". Journal of Paleontology. 96 (6): 1354–1378. doi:10.1017/jpa.2022.45. ISSN 0022-3360. S2CID 251005726.
- 1 2 3 Drovandi, Juan M.; Correa, Gustavo A.; Colombi, Carina E.; Césari, Silvia N. (2022-07-03). "Dicroidium (Zuberia) zuberi (Szajnocha) Archangelsky from exceptional Carnian leaf litters of the Ischigualasto Formation, westernmost Gondwana". Historical Biology. 34 (7): 1260–1273. doi:10.1080/08912963.2021.1974017. ISSN 0891-2963. S2CID 239437414.
- ↑ Archangelsky, S.; Brett, D. W. (1 January 1963). "Studies on Triassic Fossil Plants from Argentina: II. Michelilloa waltonii nov. gen. et spec. from the Ischigualasto Formation". Annals of Botany. 27 (1): 147–154. doi:10.1093/oxfordjournals.aob.a083828. Archived from the original on 23 August 2022. Retrieved 28 August 2022.
- ↑ Archangelsky, S.; Brett, D. W.; Harris, Thomas Maxwell (1961-07-27). "Studies on triassic fossil plants from Argentina, I. Rhexoxylon from the Ischigualasto formation". Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. 244 (706): 1–19. Bibcode:1961RSPTB.244....1A. doi:10.1098/rstb.1961.0004. S2CID 85652156. Archived from the original on 2022-08-23. Retrieved 2022-08-28.
- ↑ Martínez, L.C.A.; Artabe, A.E.; Archangelsky, S. (2020). "Studies of the leaf cuticle fine structure of Zuberia papillata (Townrow) Artabe 1990 from Hoyada de Ischigualasto (Upper Triassic), San Juan Province, Argentina". Review of Palaeobotany and Palynology. 281: 104272. doi:10.1016/j.revpalbo.2020.104272. S2CID 224943117.
Bibliography
- Geology
- Schencman, Laura Jazmín; Carina Colombi; Paula Santi Malnis, and Carlos Oscar Limarino. 2015. Diagenesis and provenance of the Los Colorados formation (Norian), Ischigualasto- Villa Unión basin, Northwest of Argentina. Revista de la Asociación Geológica Argentina 72. 219–234. Accessed 2019-03-28.
- Balabusic, Ana M., et al. 2001. Plan de Manejo del Parque Nacional Talampaya, 1–68. Administración de Parques Nacionales. Accessed 2019-03-28.
- Monetta, A.; J. Baraldo; A. Cardinali; R. Weidmann, and M. Lanzilotti. 2000. [ Distribución y características del magmatismo intratriasico de Ischigualasto, San Juan, Argentina], 644–648. IX Congreso Geológico Chileno. Accessed 2019-03-28.
- Paleontology
- Wallace, Rachel Veronica Simon. 2018. A new close mammal relative and the origin and evolution of the mammalian central nervous system (PhD thesis), 1–224. The University of Texas at Austin. Accessed 2019-03-29.
- Cabrera, Ángel. 1943. El primer hallazgo de terápsidos en la Argentina. Notas del Museo de la Plata 8. 317–331. .
Books
- Weishampel, David B.; Dodson, Peter; Osmólska, Halszka, eds. (2004). The Dinosauria, 2nd edition. Berkeley: University of California Press. ISBN 0-520-24209-2. Retrieved 2019-02-21.
Further reading
- Aceituno Cieri, P.; M.E. Zeballos; R.J. Rocca; R.D. Martino, and C. Carignano. 2015. Condicionantes geológicos en el cruce de la sierra de Valle Fértil. San Juan - Geological constraints at the crossing of sierra Valle Fertil. San Juan. Revista de Geología Aplicada a la Ingeniería y al Ambiente 35. 57–69. .
- F. E. Novas. 1986. Un probable teropodo (Saurischia) de la Formacion Ischigualasto (Triasico Superior), San Juan, Argentina [A probable theropod (Saurischia) from the Ischigualasto Formation (Upper Triassic), San Juan, Argentina]. IV Congreso Argentino de Paleontologia y Bioestratigrafia 1:1-6
- V.H. Contreras. 1981. Datos preliminares sobre un nuevo rincosaurio (Reptilia, Rhynchosauria) del Triasico Superior de Argentina. Anais II Congresso Latino-Americano Paleontologia, Porto Alegre 2:289-294
- Bonaparte, J.F. 1978. El Mesozóico de América de Sur y sus Tetrapodos - The Mesozoic of South America and its tetrapods. Opera Lilloana 26. 1–596. .
- R. M. Casamiquela. 1967. Un nuevo dinosaurio ornitisquio triasico (Pisanosaurus mertii; Ornithopoda) de la Formación Ischigualasto, Argentina [A new Triassic ornithischian dinosaur (Pisanosaurus mertii; Ornithopoda) from the Ischigualasto Formation, Argentina]. Ameghiniana 4(2):47-64
- O. A. Reig. 1963. La presencia de dinosaurios saurisquios en los "Estratos de Ischigualasto" (Mesotriasico Superior) de las provincias de San Juan y La Rioja (República Argentina) [The presence of saurischian dinosaurs in the "Ischigualasto beds" (upper Middle Triassic) of San Juan and La Rioja Provinces (Argentine Republic)]. Ameghiniana 3(1):3-20